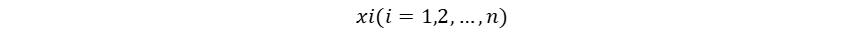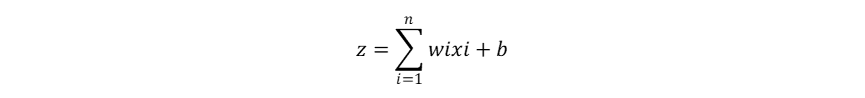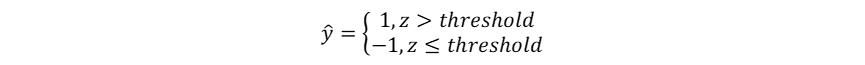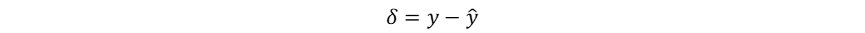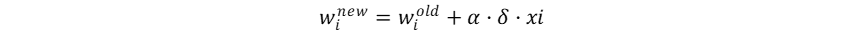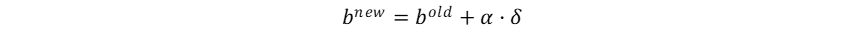Hutchinson作为现代生态学之父,其多元资源生态位理论是当前生态位理论的主流。这一理论及其应用基于一个重要的假设:物种属性决定物种适应性,物种适应性决定生态位,也就是物种属性直接决定物种生态位,而物种生态位的确定性作用决定群落物种共存和多样性动态。这一假设使得物种生态位成为了可以被量化的物种属性,但当我们将物种生态位认定为固有的物种属性,就忽视了生物适应、生物间相互作用、生物对环境的反馈在生态位形成中的影响,也暗示了生态位保守假说。
生态位概念的萌芽
生态位概念由Johnson提出[1],由Grinnell具体化[2],强调物种对环境条件的需求[3]。Elton对生态位进行了重新定义,更强调物种的功能[4]。两种概念的本质都是生物和环境的相互作用,只是侧重不同。侧重有机体(物种)对环境需求的是生境生态位,广泛应用于生物地理学研究;侧重有机体(物种)影响其他生物及环境的功能生态位,广泛应用于群落生态学研究。
在以上生态位概念的基础上,竞争排斥原理被提出,即生态位相似的物种难以稳定共存[5]。这一原理在实验中得到了证实[6],成为生态位理论/群落生态学理论的基石。自此,物种间生态位差异被用于解释物种分布、群落构建和群落动态等。
生态位概念的发展
Hutchinson对生态位概念进一步发展,认为生态位是物种所需环境条件的综合。物种属性决定物种适应性,物种适应性决定生态位,也就是物种属性直接决定物种生态位,而物种生态位的确定性作用决定群落物种共存和多样性动态[7]。其贡献在于将生态学研究从现象描述推向了假设检验,构建了通过研究物种属性研究群落物种共存和多样性动态的理论桥梁,但生态位的确定性作用作为假设,并未得到严格证明。
Hutchinson的理论将生态位分为基础生态位和现实生态位[7],两者的比较研究有助于阐明物种对生物-非生物整体环境的反馈和影响。但是无论基础生态位还是现实生态位,都无法被直接观测,只能通过观测现实生态位在真实地理空间中的投影,即物种分布状况,进行间接的不完全的观测。
生态位理论的发展
MacArthur实现了对生态位的量化,用于解释物种生态位和群落生物多样性的动态平衡规律[8]。而这些研究工作的前提是有关生态位两个基本假设,生态位保守,生态位作用的确定性。但同时MacArthur也肯定群落构建随机过程的重要性[9],只是MacArthur之后,缺少对生态位构建扩散限制和随机过程的研究。
生态位理论的巩固
Tilman的资源比例假说[10]支持了Hutchinson的多维资源生态位理论[11],解释了物种多样性的稳定机制[12],解释了生物多样性的影响[13]。
对生态位理论的质疑
生态位理论以及资源比例假说在其假设前提和适用性方面受到质疑,当代物种共存理论[14]对资源比例假说做出修正,首先考虑适合度,其次再肯定生态位的决定性作用。
而零模型[15]和群落中性理论[16]无视生物属性及其相互作用,仅考虑扩散限制、环境波动等随机过程,否定基于生物属性的生态位确定性作用。虽然从假设、逻辑推理和机理解释上存在显著的缺陷,但能够以较少的参数的预测较多的模式,迫使生态学家正视群落构建随机过程的重要性。
生态位模拟
生态位模拟的原理:受Hutchinson生态位理论的影响,主流生态学界长期使用还原论的逻辑,通过量化物种属性与环境的关系,量化物种的生态位,从而解释和预测群落动态、物种共存等现象。在生态位模拟过程中,就是通过量化物种分布与环境的关系,量化物种的生态位,从而模拟物种在当前气候情景下的潜在分布区,以及预测未来气候情景下的分布区域。
群落构建随机过程的体现:在现阶段的生态位模拟工作中,受限于生态位理论的基本假设,暂时无法体现群落构建随机过程的重要性。
生物适应、生物间相互作用、生物对环境反馈作用的体现:物种分布数据来源于真实地理空间,本身能够体现物种间的相互作用以及该物种对环境的反馈在生态位形成中的影响。
生态位保守假说的体现:生态学家无法获取真实未来的物种分布数据以及气候、土壤数据等环境数据,所以生态位模拟得到的永远只能是过去或者当前的物种生态位,当我们需要将模拟得到的生态位投影到其它时间、空间时,要求该物种生态位具有保守性。也就是说,我们希望该物种的生态位在一定的时空尺度内保持不变。
针对生态位保守假说的改进:静止是相对的,而运动是绝对的;生态位保守是相对的,而生态位变化是绝对的。生态位保守假说对于不同的研究对象,在不同的时空尺度无法绝对生效。生态位保守特征的区别,究其原因是生物属性的区别造成的生物适应特征的区别,而生物适应特征的区别广泛存在于不同层次水平。生态位保守假说作为生态位模拟的基本假设,其适用性应当在生态位模拟之初得到考量和验证。某一物种生态位保守的程度直接决定其模型的泛化能力,也就是模型投影至另一地理空间或时间下的合理性。对团队内生态位模拟工作的初步改进思路是,在生态位模拟工作起始阶段对工作的合理性做一次检验,也就是采用生态位演化速率(niche evolution rate, NER)对生态位保守的特征进行量化,暂时保留使用入侵物种生态位漂移量化方法的可能。如能够获取连续时间序列的物种分布数据,结合变化的生物气候变量进行分析,有希望实现对较短时间尺度内物种生态位变化(保守)特征的量化乃至通用指标的开发,研究对象初步定为具有扩散能力强、扩散距离远的“类入侵植物(黄帚橐吾等典型毒杂草)”。
[1] Johnson, R.H. (2009). Determinate Evolution in the Color-Pattern of the Lady-Beetles.
[2] Grinnell, J. (1917). The Niche-Relationships of the California Thrasher. The Auk, 34, 427-433.
[3] Grinnell, J. (1924). Geography and Evolution. Ecology, 5, 225-229.
[4] Elton, C.C. Animal Ecology. Nature, 119, 193-193.
[5] Colwell, R.K., & Rangel, T.F. (2009). Hutchinson’s duality: The once and future niche. Proceedings of the National Academy of Sciences, 106, 19651 – 19658.
[6] Gauze, G.F. (1934). The struggle for existence, by G. F. Gause.
[7] Hutchinson, C.E. (1957). Concluding remarks, Coldspring Harbor Symposium.
[8] Macarthur, R.H., & Levins, R.A. (1967). The Limiting Similarity, Convergence, and Divergence of Coexisting Species. The American Naturalist, 101, 377 – 385.
[9] KeChang, N., Yining, L., Zehao, S., Fangliang, H., & Jing-yun, F. (2009). Community assembly: the relative importance of neutral theory and niche theory. Biodiversity Science, 17, 579-593.
[10] Tilman, D. (1983). Resource competition and community structure. Monographs in population biology, 17, 1-296 .
[11] Tilman, D. (1988). Plant Strategies and the Dynamics and Structure of Plant Communities.
[12] Tilman, D., & Pacala, S.W. (1993). The maintenance of species richness in plant communities.
[13] Tilman, D., Lehman, C., & Thomson, K.T. (1997). Plant diversity and ecosystem productivity: theoretical considerations. Proceedings of the National Academy of Sciences of the United States of America, 94 5, 1857-61 .
[14] Chesson, P. (2000). Mechanisms of Maintenance of Species Diversity. Annual Review of Ecology, Evolution, and Systematics, 31, 343-366.
[15] Connor, E.F., & Simberloff, D. (1979). The Assembly of Species Communities: Chance or Competition? Ecology, 60, 1132-1140.
[16] Rosindell, J., Hubbell, S.P., & Etienne, R.S. (2011). The unified neutral theory of biodiversity and biogeography at age ten. Trends in ecology & evolution, 26 7, 340-8 .